
Understanding and properly using B6 for better cardiovascular health, CNS health, sleep, kidney health, brain health, immune system health, mood and much more.

It’s George.
Vitamin B6 is quite misunderstood in the general health space.
The reason for this is mainly because plenty of people accidentally, unknowingly, or whatever, consumed 50-100-150-200-300mg of pyridoxine a day for months or even years.
While vitamin forms such as methylfolate and methylcobalamin are increasingly used in supplements when it comes to B9 and B12 for example, this is not the case for B6, where pyridoxine is primarily used.
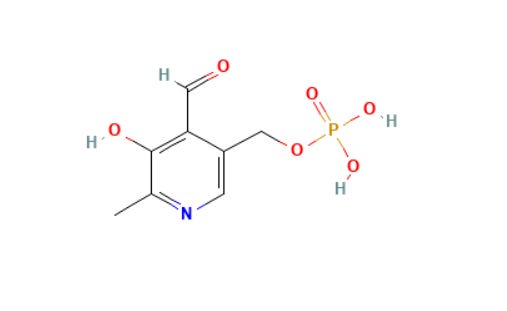
But remember that high doses of it inhibit PDXK(*), the enzyme that converts pyridoxine to P5P.
This leads to the buildup of inactive pyridoxine which is in fact neurotoxic, particularly to sensory neurons.
(*) Pyridoxine is converted to pyridoxal 5'-phosphate through phosphorylation (it is phosphorylated by the enzyme pyridoxal kinase (PDXK) to pyridoxine-5'-phosphate (PNP) a process that needs zinc and Mg) and oxidation (via pyridoxine-5'-phosphate oxidase, requiring riboflavin/FMN so it needs B2) to the active P5P.

But consuming 100mg of pyridoxine a day and saying that vitamin B6 is bad, is as ridiculous as me saying that 10-20mg of PLP per day can cure years of anxiety, depression, insomnia, ADHD symptoms so strong that the person received an actual diagnosis and do more than years of therapy and meditation.
Oh the later is true by the way for my best friend’s wife for whom low doses of PLP properly cycled have changed her life more than anything else.
Every symptom is either at remission or improved by something like 80% for 3+ years.
If you are skeptical of this:
A) You should because it’s extremely unlikely that 99.99999% of people will experience something similar
B) Let me remind you that the only essential vitamin cofactor for the enzyme glutamic acid decarboxylase is B6.
Or i am not seeing people with slow COMT saying how toxic methylated B12 is and how it can even cause anxiety attacks (to be fair, this might become a trend in a year or two # toxic B12 theory (you read about it here first)).
The point i’m trying to make with these examples, is that as humans, we are wired to pay attention to things that go wrong.
So even if horror stories are the same number as the very successful stories for a given tool, we’ll pay far more attention to the first than to the second.
Yes, B6 (especially pyridoxine) can be toxic.
But so can niacin, thiamine, riboflavin, folinic acid, zinc, manganese, molybdenum, copper and pretty much any nutrient.
In fact, sometimes, an excess creates the same symptoms as a deficiency (sometimes indirectly by depleting or harming the metabolism of other nutrients).
Both a lack of molybdenum and excess molybdenum increase the risk of gout and hyperuricemia, for example.

But to put an end to this, a toxicity usually requires at least one of the following:
Isolated supplementation of the wrong form
Supplementation of any form for too long
High doses
Certain genetic mutations
Now that we’ve got these out of the way, let’s start by breaking down why B6 is important.
Vitamin B6 includes the family of pyridoxal, pyridoxine (pyridoxol), pyridoxamine, and their phosphorylated derivatives.
The body cannot synthesize them de novo (in meaningful amounts) and must obtain them from diet or supplements.
The bioactive coenzyme form is pyridoxal 5’-phosphate (PLP or P5P), which participates in over 100 enzymatic reactions, accounting for roughly 4% of all human enzymatic activity whether that’s called protein (amino acid) metabolism, neurotransmitter production, hemoglobin synthesis, and one-carbon metabolism.
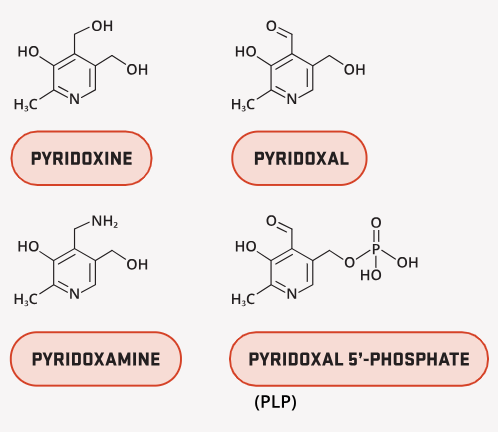
Some of its key functions include:
Nervous system “regulation”
PLP serves as a critical coenzyme for multiple enzymes involved in neurotransmitter biosynthesis, making vitamin B6 essential for proper brain signaling, mood regulation, cognition, and stress response.
Key reactions include:
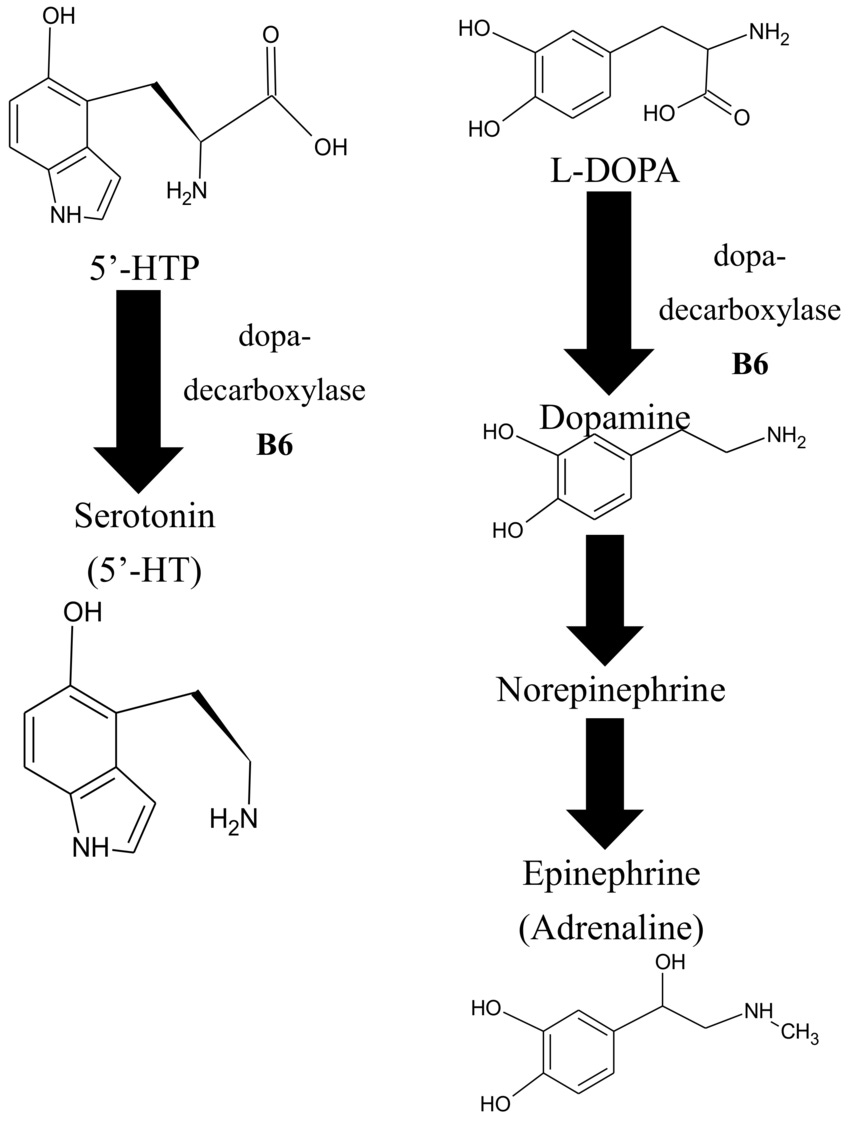
Serotonin synthesis: The PLP-dependent enzyme aromatic L-amino acid decarboxylase (AADC) catalyzes the final step, converting 5-hydroxytryptophan (5-HTP, derived from tryptophan) into serotonin (5-HT).
Dopamine (and catecholamines): The same AADC enzyme decarboxylates L-DOPA to dopamine, downstream steps also support norepinephrine and epinephrine.
GABA production: Glutamate decarboxylase (GAD), with two main isoforms, GAD65 (synaptic, activity-dependent) and GAD67 (cytosolic, basal levels), requires PLP as a cofactor to convert excitatory glutamate into inhibitory GABA. This is one of the most PLP-sensitive reactions in the brain. GAD65 exists largely as an apo-enzyme (inactive without PLP), so PLP availability directly modulates GABA production capacity.
Additional neurotransmitters and modulators:
Glycine: Via serine hydroxymethyltransferase (SHMT).
D-serine: An important NMDA receptor co-agonist.
Glutamate: Via transaminases.
Histamine: Via histidine decarboxylase.
So adequate PLP helps maintain balance between excitatory and inhibitory signaling.
This is why suboptimal status or genetic variants impairing B6 activation/conversion such as PDXK, PNPO, or ALPL, can contribute to hyperexcitability, anxiety, racing thoughts (hypomania-like states), insomnia, ADHD traits, amygdala overactivity, and mood instability overall.
Hemoglobin synthesis and function
PLP is the essential cofactor for δ-aminolevulinic acid synthase (ALAS), specifically the erythroid-specific isoform ALAS2 in bone marrow precursors, which catalyzes the rate-limiting first step of heme biosynthesis.
The reaction condenses glycine and succinyl-CoA (from the mitochondrial TCA cycle) to form δ-aminolevulinic acid (ALA) in the mitochondria.
PLP binds to a conserved lysine residue in ALAS2, stabilizing the catalytic site and enabling Schiff base formation with glycine.
Subsequent steps occur in both mitochondria and cytosol, ultimately producing protoporphyrin IX, which incorporates ferrous iron (Fe²⁺) via ferrochelatase to form heme.
Heme is then incorporated into globin chains to produce hemoglobin (in erythrocytes) or myoglobin (in muscle).
So consequences of impaired PLP availability include:
Reduced ALAS2 activity → decreased heme production.
Iron continues to enter mitochondria but cannot be properly utilized, leading to mitochondrial iron overload.
Characteristic ringed sideroblasts (erythroblasts with iron-laden mitochondria forming a ring around the nucleus).
Resulting anemia is classically sideroblastic (often microcytic and hypochromic due to insufficient hemoglobin), though recent studies show it can also present as normocytic or even macrocytic in some cases of acquired deficiency.
Vitamin B6-responsive sideroblastic anemia (congenital or acquired) often improves dramatically with pyridoxine or P5P supplementation, as it restores ALAS2 function.
Beyond synthesis, PLP and pyridoxal can also bind covalently or non-covalently to hemoglobin.
This interaction modestly decreases hemoglobin’s affinity for oxygen (right-shift in the oxygen dissociation curve), facilitating easier oxygen unloading to tissues.
This allosteric effect is similar to (but weaker than) that of 2,3-bisphosphoglycerate (2,3-BPG).
While demonstrable in vitro and in supplemented red cells, its physiological significance in healthy individuals at normal B6 levels remains unclear and is considered minor.
Regulating the kynurenine pathway
Approximately 90–95% of dietary tryptophan (beyond what is used for protein synthesis and serotonin production) is metabolized through the kynurenine pathway (KP), the primary de novo route for synthesizing nicotinamide adenine dinucleotide (NAD+), a critical coenzyme for energy metabolism, DNA repair, sirtuins, and over 500 cellular reactions.
The pathway begins with the rate-limiting enzymes indoleamine 2,3-dioxygenase (IDO1/IDO2) or tryptophan 2,3-dioxygenase (TDO), which convert tryptophan to N-formylkynurenine (then kynurenine).
PLP plays a pivotal role further downstream as the essential cofactor for two key enzymes: kynureninase (KYNU) and kynurenine aminotransferase (KAT).
In case you missed it, read this:

Now (read the article first for the following to make sense), here are some critical PLP-dependent steps:
Kynureninase (KYNU): Requires PLP to hydrolyze
Kynurenine → anthranilic acid
3-Hydroxykynurenine (3-HK) → 3-hydroxyanthranilic acid (a key step toward quinolinic acid → NAD+).
Kynurenine aminotransferase (KAT): Also PLP-dependent; produces kynurenic acid (KYNA) from kynurenine and xanthurenic acid (XA) from 3-HK.
So when PLP availability is low kynureninase activity is particularly impaired because it has a relatively high Km for PLP.
This leads to:
Accumulation of upstream metabolites:
Kynurenine
3-Hydroxykynurenine (3-HK, pro-oxidant, generates reactive oxygen species)
Xanthurenic acid (XA)
Kynurenic acid (KYNA) in some branches
Impaired NAD+ production: Reduced flux through the pathway limits formation of quinolinic acid and subsequent NAD+.
This affects cellular energy (ATP via mitochondria), DNA repair (PARP enzymes), epigenetic regulation (sirtuins), and redox balance.
Shift toward neuroactive or pro-inflammatory metabolites:
Quinolinic acid (QUIN): NMDA receptor agonist → excitotoxicity, oxidative stress, and neuroinflammation (implicated in depression, neurodegeneration, and cognitive issues).
3-HK: Promotes apoptosis and free radical damage.
KYNA: Generally neuroprotective (NMDA antagonist) but in excess can impair glutamatergic signaling.
Overall imbalance favors inflammation and neuronal stress.
nature.com
The KP is strongly induced during immune activation and chronic inflammation:
Cytokines (especially IFN-γ) upregulate IDO1, increasing tryptophan catabolism.
This creates a feedback loop where inflammation depletes PLP (via increased demand and oxidative degradation), further impairing the pathway and amplifying metabolite imbalances.
Chronic low-grade inflammation thus raises vitamin B6 requirements and lowers plasma PLP levels.
This mechanism links suboptimal B6 status to:
Cardiovascular disease (via endothelial dysfunction and oxidative stress)
Depression (reduced serotonin + neurotoxic metabolites)
Neurodegenerative conditions
Insulin resistance/diabetes (XA and certain metabolites interfere with insulin action)
Immune dysregulation (altered T-cell responses and tolerance)
*There are studies at the end.